
Potential mechanism of NO participating in melatonin alleviating the postharvest browning of lotus seeds
-
摘要:
为研究NO参与褪黑素(melatonin,MT)延缓莲子采后褐变的作用机理,该研究采用蒸馏水、NO、MT、一氧化氮合酶抑制剂(nomega-nitro-l-arginine methyl ester,L-NAME)、硝酸还原酶抑制剂(tungstate,TUN)和NO清除剂(Carboxy-PTIO,cPTIO)、MT + L-NAME、MT + TUN和MT + cPTIO对莲子进行处理。MT处理的莲子内源NO含量显著高于对照组(P < 0.05),且一氧化氮合酶(nitric oxide synthase,NOS)活性在贮藏前3 d提高60.36%~71.08%,NO合成途径中关键物质L-精氨酸和瓜氨酸含量显著高于对照组(P < 0.05),尤其在第1天时瓜氨酸含量为对照组的1.44~1.59倍,但对硝酸还原酶(nitrate reductase,NR)活性的影响并无规律;当MT结合NOS抑制剂L-NAME对莲子处理时,莲子内源NO含量显著低于对照组(P < 0.05),且MT保鲜效果消失;当结合NR抑制剂TUN时,莲子内源NO含量显著高于对照组(P < 0.05),且MT仍具有保鲜效果,因此初步确定MT通过NOS途径诱导莲子内源NO生成。在此基础上,通过分析鉴定得出莲子中主要酚类物质为儿茶素(占总酚含量60%以上),后续重点分析了MT通过NO调控莲子儿茶素代谢的机理。结果得出,MT处理的莲子多酚氧化酶(polyphenol oxidase, PPO)活性低于对照组35.39%~57.36%,同时该处理显著提高了其儿茶素合成代谢关键酶肉桂酸-4-羟化酶(cinnamate-4-hydroxylase,C4H)、二氢黄酮醇还原酶(dihydroflavonol 4-reductase,DFR)、查尔酮合成酶(chalcone synthase,CHS)、查尔酮异构酶(chalcone isomerase,CHI)和植物无色花青素还原酶(colorless anthocyanin reductase,LAR)活性(P < 0.05),由此其儿茶素含量在1 d后高于对照组15.35%~47.86%,但当MT结合L-NAME或cPTIO处理时,该效果消失。因此,外源MT通过NOS途径诱导NO合成造成NO的大量积累,并作用于莲子儿茶素,一方面抑制其与PPO产生酶促褐变,另一方面促进其合成,最终延缓莲子采后褐变进程。
Abstract:This study aims to investigate the potential mechanism of melatonin (MT) to alleviate the browning of postharvest lotus seeds. 1) The samples were treated with the distilled water, nitric oxide (NO), MT, the inhibitor of nitric oxide synthase (NOS) nomega-nitro-L-arginine methyl ester (L-NAME), the inhibitor of nitrate reductase (NR) tungstate (TUN), and the scavenger of NO carboxy-PTIO (cPTIO), MT + L-NAME, MT + TUN and MT + cPTIO. A systematic analysis was implemented to explore the possible pathway of NO biosynthesis in the lotus seeds. Then, the levels of key enzymes and substances were determined to involve in the NO biosynthesis. The results showed that the endogenous NO content of the MT treated lotus seeds was significantly higher than that of the control. The NOS activity of lotus seeds treated with MT was promoted by 60.36%−71.08% before 3 days of storage. In addition, the MT treatment significantly increased the contents of L-arginine and citrulline (P <0.05), which were the key substances in the NO biosynthesis. It was notable that the citrulline content in the MT treated sample was 1.44~1.59 times than those of the controls. By contrast, there was the varying influence of MT treatment on the activity of NR in lotus seeds. There was the beneficial effect of MT on the lotus seeds and the endogenous NO content of the tissue, when the MT was combined with the inhibitor of NR (TUN). However, this beneficial effect disappeared, when the MT was combined with the inhibitor of NOS (L-NAME) for treating the lotus seeds. The endogenous NO content in this treatment was significantly lower than that in the control. Therefore, the exogenous MT treatment was induced the NO biosynthesis in lotus seeds through the pathway of NOS. 2) The potential mechanism of MT was clarified to regulate the browning of lotus seeds. The phenolic compounds were identified in the lotus seeds using a LC20 HPLC (Shimadzu, Japan) system coupled to a TripleTOF® 5600 + quadrupole time-of-flight (QTOF) mass spectrometer equipped with a DuoSpray™ ion source (Sciex, Ontario, Canada). It was found that there were six flavanols and seven phenolic acids in the lotus seeds, and the content of catechin, a type of flavanol, accounts for over 60% of the total phenol content, the following was ellagitannin, a type of phenolic acid, accounts for 7.45% of the total phenol content. It infers that the catechin was the major phenolic compound of lotus seeds. The flavan-3-ol was the most common direct natural substrate of polyphenol oxidase (PPO) in the plant, especially for the catechin and epicatechin. Thereby, the potential mechanism of MT was determined, where the catechin metabolism was regulated to induce the NO production in the following experiments. The results indicated that the activity of PPO in the MT treated lotus seeds was lower by 35.39%−57.36% than those in the control. What’s more, the MT treatment significantly increased the activities of key enzymes, including cinnamate-4-hydroxylase, dihydroflavonol reductase, chalcone synthase, chalcone isomerase and colorless anthocyanin reductase (P <0.05) in the catechin synthetic metabolism. As a result, the content of catechin in the MT treated sample was higher by 15.35%−47.86% than those in the control after 1 days of storage. Whereas, this positive effect was negated, when the MT was combined with L-NAME or cPTIO for treating lotus seeds. Therefore, it was concluded that the exogenous MT treatment was induced the biosynthesis of NO through NOS pathway, and then the accumulated NO acted on the catechin metabolism of lotus seeds. The MT treatment was used to inhibit the PPO activity, and then the catechin was suppressed to participate in the enzymatic browning; The biosynthesis of catechin was promoted in this treatment. Consequently, the lotus seeds browning was also alleviated by the MT treatment. These findings can provide the theoretical and technical support to preservation of lotus seeds and the signal transduction between MT and NO.
-
Keywords:
- lotus seeds /
- melatonin /
- nitric oxide /
- browning /
- catechin metabolism
-
0. 引 言
光伏农业系统是将太阳能发电与农业生产有机耦合的新型发展模式。随着可再生能源的推广和农业现代化的需求,该模式在全球范围内得到了快速发展[1-2]。2021年的国际光伏农业大会指出,中国已成为光伏农业模式发展最好的国家之一,农光一体总装机容量位居世界第一[3]。目前,光伏农业项目在中国3种类别的太阳能资源区均有应用推广,为光伏产业发展和乡村振兴作出了重要贡献[4-5]。
在农业用地上建设的光伏阵列是光伏农业系统的主体结构,其主要由光伏组件与支撑部件等组成,具体包括固定支架、柔性支架与跟踪式支架3种光伏阵列类型[6-7]。鉴于施工成本及设备运行的可靠性,目前绝大部分的光伏农业项目会优先选用固定支架的安装方案[6]。为充分保障光伏农业系统中农业生产的土地利用率和机械化率,中国江苏、云南等省份相继发文要求光伏农业项目的光伏组件在具有一定的倾角下,设计足够的铺设高度以保证农业的正常生产。通过调研发现,截至2024年6月,已有江苏、陕西等13个省份对光伏农业系统中光伏组件下沿边缘距离地面的垂直高度(以下简称光伏组件铺设高度)做了强制性规定,其中江苏等大多省份的光伏农业项目最低光伏组件铺设高度为2.5 m。
虽然最低光伏组件铺设高度在一定程度上保障了农业的生产,但是仍有大量的光伏农业项目在实际生产过程中存在“重光轻农”的现象[8-9]。在光伏农业项目规划建设时,铺设的大量光伏组件会产生较大的遮阴面积与动态阴影带,从而影响光伏农业系统内部的光环境,最终影响农作物的产量,这在一定程度上限制了光伏农业产业的发展[10-12]。针对上述情况,国内外学者对光伏农业系统内部的光环境进行了深入研究。西北农林科技大学邹志荣团队以光伏日光温室为研究对象,探究了光伏组件遮阴对室内光环境及番茄生长的影响[13];浙江大学郑荣进团队分析了光伏茶园中固定支架与跟踪式支架的光伏组件遮阴对茶园内部光环境及茶树生长的影响[14];SANTRA等[15]以3种光伏组件铺设密度的光伏阵列为研究对象,研究了光伏组件遮阴对3种光伏农业系统内部光量子通量密度的影响;江苏省农业科学院鲍恩财团队结合传感器配置策略和光伏农业系统空间结构特征,揭示了光伏组件铺设高度为3.2 m的光伏农业系统内部光环境在夏、冬两季的变化规律[16-17]。
上述研究有效表征了光伏农业系统内部的光环境特征,不过未考虑光伏组件铺设高度这一结构参数对其内部太阳辐射强度的变化的影响。另外,光伏组件在光伏农业系统内产生的阴影带随时间变化而变化,关于光伏农业系统中光伏组件遮阴宽度的动态变化规律亦未见报道。目前对农业建筑中阴影动态变化规律的研究仅局限在连栋温室天沟[18]、日光温室固定保温被 [19]等结构。
已有研究发现光伏组件遮阴对甘薯[20]、小麦[21]、蔬菜[22]及猕猴桃[23]等作物的产量均会造成负面影响,而无花果(Ficus carica L.)是一种适应性强、耐旱、耐荫、挂果早、产量高、抗病性好且栽植容易的经济性苗木,在江苏、云南、陕西和湖南等地均有种植,能较好适应光伏农业系统内部的复杂环境[24-25]。故本研究以栽植无花果树的光伏农业系统为研究对象,通过试验测试分析光伏组件铺设高度对光伏农业系统内部太阳辐射强度的影响。根据太阳直射辐射理论计算光伏组件遮阴宽度,并对受不同光伏组件铺设高度和地理位置影响而形成的阴影宽度进行分析。同时从光伏组件遮阴的角度定量分析光伏组件铺设高度对无花果产量和叶片光合特性的影响。
1. 材料与方法
1.1 试验光伏农业系统描述
供试光伏农业系统位于江苏省南京市溧水区的光伏农业园区(31.62°N,119.18°E)。该园区于2016年建成,占地约47 hm2,包含8个光伏阵列片区,总装机容量为20 MW,每年可发电约2 400万kW·h。供试光伏农业阵列(以下简称光伏阵列)坐北朝南,跨度为6.8 m,采用横向4排光伏组件紧密排布方式,如图1所示。光伏组件的铺设高度均不低于2.5 m,垂直投影比例(光伏组件在农田上的垂直投影总面积/架设光伏组件的农田面积)为53.3%,由混凝土预制桩等结构支撑,安装倾角为24°,材料为265 Wp多晶硅,光电转换效率为16.3%。单块光伏组件尺寸为
1640 mm×992 mm×35 mm。为保证光伏农业系统内部的采光率和机械化作业空间,目前江苏省现存光伏农业项目的光伏组件铺设高度区间为2.5~4.0 m。基于此,本研究选取了光伏组件铺设高度为2.5、3.2、3.9 m的3种光伏农业系统。除光伏组件铺设高度外,供试光伏农业系统的其他结构和材料参数完全一致。
1.2 栽植果树
2019年初在供试光伏阵列内部南北向居中位置,沿东西向栽植了单行矮化无花果(品种:青皮),如图1a所示。无花果树株间距约3.0 m,株高约1.0 m,冠幅约2.5 m,栽植密度约490株/hm2,每年9—10月为采收期。
1.3 试验设计
1.3.1 种植区域光环境
结合无花果株高等生长参数,距地面1.0 m高度的太阳辐射强度大小及分布情况是评价光伏农业系统内部光环境的重要指标。基于此,本文监测的光环境因子包括2.5、3.2、3.9 m的光伏组件铺设高度下(分别对应为H2.5、H3.2、H3.9)光伏农业系统内部的太阳辐射强度,其中分为光伏阵列板下种植区域(U)和板间种植区域(B)2个部分。同时,在系统外部开阔地设置气象站,同步监测露天(CK)的太阳辐射强度,具体种植区域分类及测点布置如图2所示。本文以单跨光伏阵列北侧板下种植区域的太阳辐射强度测点数据作为评价光伏阵列板下种植区域太阳辐射强度的数据来源。试验时间为2023年无花果的全生育期,即2023年4月1日—2023年9月30日。
![]() 图 2 试验测点布置图注:H 为光伏组件铺设高度,m。Figure 2. Layout of test pointsNote: H represents the vertical distance between the lower edge of the photovoltaic module and the ground, m.
图 2 试验测点布置图注:H 为光伏组件铺设高度,m。Figure 2. Layout of test pointsNote: H represents the vertical distance between the lower edge of the photovoltaic module and the ground, m.测试仪器采用HOBO系列传感器。所有传感器进行全天监测,数据记录时间间隔为10 min。研究所用的太阳辐射强度传感器为太阳总辐射传感器(S-LIB-M003,Onset Computer Crop公司,美国,精度为±10 W/m2),由HOBO微型气象站(H21-USB,Onset Computer Crop公司,美国)采集。钟坤炎等 [26] 在研究全年太阳辐射规律时,明确中国夏半年地方时06:00前为太阳升起时段、18:00后为太阳落下时段。基于此,本研究定义夏季白昼时间段为06:00—18:00,夜间时间段为18:00至次日06:00 。
采光率作为光环境的重要评价指标,在各类建筑的光环境分析中都有应用。张勇等[27]将日光温室内测点光照度均值与室外测点光照度均值的比值作为透光率来评价日光温室采光性能。基于此,本文结合光环境试验,规定以光伏农业系统内部测点的太阳辐射强度平均值与外部太阳辐射强度平均值的比值作为该测点的采光率,以某一测点在多个时间段下采光率的均值作为该测点的平均采光率,将采光率和平均采光率作为光伏农业系统内部光环境的评价指标,并通过式(1)~(2)计算。
Rd=1kk∑k=1Eink/1kk∑k=1Eoutk (1) Ravg=1jj∑j=1Rdj (2) 式中Ein为内部太阳辐射强度的测量值,W/m2;Eout为外部太阳辐射强度的测量值,W/m2;Rd为试验期间日采光率,%;Ravg为试验期间平均采光率,%;k为太阳辐射强度数据日记录条数;j为试验天数,d。
1.3.2 作物产量、光合特性
为探究光伏组件遮阴对作物产量和光合特性的影响,本研究对光伏农业系统内部无花果产量和叶片光合特性进行测定分析。本试验以光伏组件铺设高度为试验因素,设置H2.5、H3.2、H3.9共3种处理,以露天种植为对照(CK),在每个高度下选取5棵生长状况良好、具有代表性的果树进行挂牌。试验的所有处理在同一块大田,氮、磷、钾肥用量相同。试验期间除施肥外,不涉及其他农艺措施。
在产量对比试验中,于9月份果实成熟期对每个处理的挂牌果树进行一次性采收并统计果数。同时在每个处理下各选取果树外围中部长势一致且无病虫害的成熟果实15个,并现场进行单果质量等产量构成指标测定。
在光合参数对比试验中,于9月份果实成熟期使用LI-COR 6800型便携式光合仪(LI-COR公司,美国)测定无花果树冠面中间位置叶片的光合特性,光合仪参数设定为环境温度25 ℃,相对湿度为50%,光量子通量强度为1 000 μmol/(m2·s),CO2浓度为400 μmol/mol,测定系统采用开放式气路,自然光源;每个处理选取长势一致、生长良好的3棵果树,重复3次,取均值;测定的叶片光合参数包括净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)。
1.4 数据处理
光伏组件铺设高度对无花果产量及叶片光合特性的影响采用单因素方差分析,差异显著性检验采用Duncan新复极差法。根据云量的观测情况和日照时数,将试验期间的天气类型划分为晴天、阴天和雨天[28]。采用Microsoft Excel 2010软件对试验数据进行计算和绘图,采用SPSS 20统计软件进行统计分析。
2. 光伏组件遮阴宽度的确定方法
2.1 光伏组件遮阴宽度定义
为研究光伏组件在光伏农业系统内形成的阴影宽度,现将遮阴宽度d进行如下定义:单侧光伏组件在光伏阵列间隔处地面上的投影宽度(OP),如图3所示。
![]() 图 3 光伏农业系统内部遮阴宽度的形成注:A为光伏组件东北侧顶点与太阳光线的交汇点;B为太阳光线透过A点在地面上的投影点;O为A点在地面上的垂直投影点;P为B点在Y轴上的垂直投影点;W为单侧光伏组件的宽度,m;β为光伏组件的倾角,(°);γs为太阳方位角,(°);h为太阳高度角,(°)。Figure 3. Formation of internal shading width in an agrivoltaic systemNote:A represents the point where direct sunlight meet the northeast vertex of the photovoltaic (PV) panel; B represents projection point on the ground of direct sunlight through point A; O represents vertical projection point on the ground of point A; P represents vertical projection point on the Y-axis of point B; W represents the width of the single-side PV module, m; β represents the inclination angle of the PV panel, (°); γs represents the azimuth of the sun, (°); h represents the altitude angle of the sun, (°).
图 3 光伏农业系统内部遮阴宽度的形成注:A为光伏组件东北侧顶点与太阳光线的交汇点;B为太阳光线透过A点在地面上的投影点;O为A点在地面上的垂直投影点;P为B点在Y轴上的垂直投影点;W为单侧光伏组件的宽度,m;β为光伏组件的倾角,(°);γs为太阳方位角,(°);h为太阳高度角,(°)。Figure 3. Formation of internal shading width in an agrivoltaic systemNote:A represents the point where direct sunlight meet the northeast vertex of the photovoltaic (PV) panel; B represents projection point on the ground of direct sunlight through point A; O represents vertical projection point on the ground of point A; P represents vertical projection point on the Y-axis of point B; W represents the width of the single-side PV module, m; β represents the inclination angle of the PV panel, (°); γs represents the azimuth of the sun, (°); h represents the altitude angle of the sun, (°).为明确受光伏组件尺寸和铺设高度等因素影响的光伏组件遮阴宽度计算方法,提出如下假设:1)光伏组件只对直射光产生阻挡,散射光在阵列内部任一个方向上具有相同的辐射强度;2)两侧光伏组件之间种植区同一时间内只受单侧或双侧光伏组件遮阴的影响;3)阵列尺寸、光伏组件数量不受限制,光伏阵列长度在东西方向上无限延伸;4)光伏组件的结构、材料一致且透光率默认为0;5)光伏组件上下对称截面为宽度忽略不计的矩形;6)光伏阵列的垂直立剖面与正南方向的夹角(方位角)默认为0。
2.2 光伏组件遮阴宽度计算方法
太阳高度角h是指太阳直射光线入射方向与地平线的夹角,h可由式(3)计算[29]。
sinh=sinη⋅sinδ+cosη⋅cosδ⋅cosω (3) 式中 η 为当地北纬纬度,(°);δ 为太阳赤纬角,(°);ω 为太阳时角,(°)。
太阳赤纬角 δ 可由式(4)计算[30]。
δ=23.45°sin(360°×284 + N365) (4) 式中N为日序数,当日期是1月1日时,N=1(1≤N≤365)。
时间角度 ω 可由式(5)计算[30]。
ω=15(Tr−12) (5) 式中Tr为真太阳时,Tr可由式(6)计算求得。
Tr=t−120−θ15+e60 (6) 式中t为北京时间,θ 为当地经度,(°),时差e可由式(7)计算。
e≈9.87sin(2A)−7.53cosA−1.5sinA (7) 式中A为常数,A可由式(8)计算求得。
A=360(N−81)364 (8) 太阳方位角 γs 是指太阳光线在地平面上的投影与当地子午线的夹角,γs可由式(9)计算[29]。
cosγs=sinh⋅sinη−sinδcosh⋅cosη (9) 为便于计算光伏组件在光伏农业系统内形成的阴影宽度,本研究以光伏组件东北角边缘点在地面上的垂直投影点作为坐标轴原点建立了三维坐标系,如图3所示。光伏阵列遮阴阴影的形成过程为:某时刻,太阳直射光线以平行于AP的角度入射至系统内部,经光伏组件遮挡后在地面形成实际阴影宽度OB,其在南北方向的投影长度OP为光伏组件在两排光伏阵列间隔处遮阴的宽度(Ls)。根据太阳位置参数、地理位置参数和几何关系,Ls可由式(10)计算求得。
Ls=(W⋅sinβ+H)⋅coth⋅cosγs (10) 3. 结果与分析
3.1 光热环境特性分析
3.1.1 太阳辐射强度
图4显示了3个光伏组件铺设高度下光伏农业系统内日平均太阳辐射与采光率的变化。H2.5、H3.2和H3.9处理下,光伏阵列板间与板下2个种植区域日平均太阳辐射强度变化范围分别为51.6~432.7、28.4~82.3 W/m2,47.6~439.2、33.4~108.6 W/m2,40.3~448.5、34.9~113.5 W/m2;平均采光率分别为:68.9%、21.2%(H2.5),65.2%、24.1%(H3.2),55.4%、26.3%(H3.9)。由图4可见,在3种光伏组件铺设高度下,光伏阵列板间种植区域的平均太阳辐射强度和采光率随着光伏组件铺设高度的增加而降低,光伏阵列板下种植区域相反。
![]() 图 4 光伏农业系统内部太阳辐射强度变化注:H2.5、H3.2、H3.9分别代表2.5、3.2、3.9 m的光伏组件铺设高度;B为光伏阵列板间种植区域,U为光伏阵列板下种植区域。Figure 4. Variation of solar radiation intensity in agrivoltaic systemsNote: H2.5, H3.2 and H3.9 represent the vertical heights of the lower edge of the PV module from the ground which are 2.5, 3.2 and 3.9 m respectively. B represents the planting area between PV modules, U represents the planting area under PV modules.
图 4 光伏农业系统内部太阳辐射强度变化注:H2.5、H3.2、H3.9分别代表2.5、3.2、3.9 m的光伏组件铺设高度;B为光伏阵列板间种植区域,U为光伏阵列板下种植区域。Figure 4. Variation of solar radiation intensity in agrivoltaic systemsNote: H2.5, H3.2 and H3.9 represent the vertical heights of the lower edge of the PV module from the ground which are 2.5, 3.2 and 3.9 m respectively. B represents the planting area between PV modules, U represents the planting area under PV modules.为深入了解不同光伏组件铺设高度的光伏农业系统内部光环境变化特征,本研究选择晴天(7月12日)、雨天(7月13日)和阴天(7月14日)的太阳辐射强度数据进行分析,其逐时变化特征如图5所示。数据显示,3种天气条件下各测点曲线具有较高的一致性,均呈现先上升后稳定波动达到峰值,最后下降变为0的变化趋势,且各曲线峰值对应的时间节点基本一致。所有测点的太阳辐射强度在12:00—14:00来回波动达到最大值。
![]() 图 5 不同天气条件下不同光伏组件铺设高度的光伏农业系统内部太阳辐射强度变化Figure 5. Variation of solar radiation intensity in agrivoltaic systems with different PV panel heights under different weather conditions
图 5 不同天气条件下不同光伏组件铺设高度的光伏农业系统内部太阳辐射强度变化Figure 5. Variation of solar radiation intensity in agrivoltaic systems with different PV panel heights under different weather conditions晴天条件下,CK日平均太阳辐射强度为477.9 W/m2,H2.5、H3.2、H3.9的光伏阵列板间和板下种植区域日平均太阳辐射强度分别为336.8、75.8 W/m2,323.3、84.2 W/m2,315.4、90.5 W/m2。由此可见,晴天条件下3种光伏组件铺设高度的光伏阵列板下和板间种植区域采光率变化区间分别为21.5%~24.7%、66.1%~70.5%,故光伏阵列板间种植区域平均采光率比整体试验期间略高,光伏阵列板下种植区域相反。
3.2 光伏组件遮阴宽度计算
由式(10)可知,遮阴宽度会受到光伏组件宽度、光伏组件倾角、光伏组件铺设高度、太阳高度角和太阳方位角5个参数的影响。其中,太阳方位角、太阳高度角受到光照时间、所在地理位置的影响,光伏组件宽度和光伏组件倾角因发电效率、发电容量等客观原因在同一地区任一光伏农业项目上基本是固定的,而光伏组件铺设高度会因地形及防洪标高一致性等因素存在差异。基于此,本文从不同的地理位置和光伏组件铺设高度这两个参数入手,计算分析其对光伏农业系统内光伏组件遮阴宽度的影响。为了量化光伏组件遮阴的宽度,本文在计算时间范围上,取1个季度为1个时间跨度,共4个跨度。
3.2.1 地理位置对遮阴宽度的影响
不同地理位置的经纬度不同,其中,经度会影响当地真太阳,纬度会影响当地太阳高度角,从而导致遮阴宽度存在差异。由于无花果在江苏、云南、陕西和湖南等省份均有大面积种植,且4个省份的最低光伏组件铺设高度均为2.5 m,故选择江苏、云南、陕西和湖南4个省份,并以4个省份的省会城市为对象分析地理位置对遮阴宽度的影响。暂不考虑光伏组件铺设高度造成的影响,即默认光伏组件铺设高度为2.5 m,且不同地理位置的光伏阵列尺寸与试验光伏阵列相同。上述地区的光伏组件遮阴宽度在整点时刻的平均值如表1所示。由表1可见,光伏组件周年遮阴宽度整体呈现先下降再上升的变化趋势,变化差异明显。在南京,无花果生育期内(4—9月)光伏组件遮阴宽度均值为0.81 m,最大值出现在中午12:00,下午遮阴宽度均值较上午大0.13 m;在昆明、西安和长沙,无花果生育期内(4—9月)光伏组件遮阴宽度均值分别为0.52、0.88和0.65 m,最大值均出现在13:00,下午遮阴宽度均值较上午分别大1.17、1.24和0.69 m。同时结合4地地理位置可以发现,同一时刻下不同地理位置的光伏组件遮阴宽度差异较大,且在无花果全生育期内光伏组件遮阴宽度随纬度增大而增大。
表 1 不同光伏组件铺设高度与地理位置对遮阴宽度的影响Table 1. Effects of photovoltaic panels with different laying heights and geographical location on the shading width月份
Months时刻Time 遮阴宽度 Shading width/m 南京Nanjing
(31.62°N,119.18°E)昆明Kunming
(25.22°N,103.10°E)西安Xi′an
(34.10°N,108.40°E)长沙Changsha
(27.90°N,112.53°E)H2.5 H3.2 H3.9 H2.5 H3.2 H3.9 H2.5 H3.2 H3.9 H2.5 H3.2 H3.9 1—3 09:00 3.76 4.40 5.04 4.82 5.64 6.47 4.98 5.83 6.67 3.93 4.60 5.27 10:00 3.24 3.79 4.34 3.38 3.95 4.53 3.68 4.31 4.94 3.20 3.74 4.29 11:00 3.02 3.54 4.05 2.91 3.40 3.90 3.25 3.81 4.36 2.91 3.41 3.90 12:00 2.95 3.45 3.95 2.71 3.17 3.63 3.08 3.60 4.13 2.79 3.27 3.74 13:00 2.97 3.48 3.99 2.64 3.08 3.53 3.03 3.55 4.06 2.78 3.25 3.72 14:00 3.11 3.64 4.17 2.65 3.10 3.55 3.09 3.61 4.14 2.85 3.34 3.83 15:00 3.46 4.05 4.64 2.77 3.24 3.71 3.28 3.84 4.40 3.07 3.59 4.11 平均 3.22 3.76 4.31 3.13 3.65 4.19 3.48 4.08 4.67 3.08 3.60 4.12 4—6 09:00 0.42 0.49 0.56 0.11 0.13 0.15 0.33 0.38 0.44 0.24 0.28 0.32 10:00 0.63 0.73 0.84 0.23 0.27 0.31 0.59 0.69 0.79 0.37 0.44 0.50 11:00 0.73 0.86 0.98 0.33 0.39 0.45 0.76 0.89 1.02 0.53 0.62 0.71 12:00 0.77 0.90 1.03 0.43 0.50 0.57 0.84 0.98 1.13 0.60 0.70 0.80 13:00 0.74 0.86 0.99 0.47 0.55 0.63 0.86 1.00 1.15 0.59 0.70 0.80 14:00 0.64 0.75 0.85 0.44 0.52 0.59 0.81 0.95 1.09 0.53 0.61 0.70 15:00 0.43 0.51 0.58 0.34 0.40 0.45 0.69 0.81 0.93 0.37 0.44 0.50 平均 0.62 0.73 0.83 0.34 0.39 0.45 0.70 0.81 0.94 0.46 0.54 0.62 7—9 09:00 0.83 0.97 1.12 0.47 0.55 0.64 0.75 0.87 1.00 0.62 0.73 0.83 10:00 0.99 1.16 1.33 0.58 0.68 0.78 0.97 1.13 1.30 0.76 0.89 1.02 11:00 1.07 1.25 1.44 0.69 0.81 0.92 1.10 1.28 1.47 0.88 1.03 1.17 12:00 1.10 1.29 1.48 0.77 0.90 1.04 1.16 1.36 1.55 0.93 1.09 1.24 13:00 1.08 1.27 1.45 0.81 0.94 1.08 1.17 1.37 1.57 0.93 1.09 1.25 14:00 1.02 1.19 1.37 0.79 0.93 1.06 1.14 1.34 1.53 0.89 1.04 1.19 15:00 0.89 1.04 1.19 0.73 0.85 0.97 1.07 1.25 1.43 0.78 0.91 1.05 平均 1.00 1.17 1.34 0.69 0.81 0.93 1.05 1.23 1.41 0.83 0.97 1.11 10—12 09:00 4.49 5.26 6.02 5.62 6.57 7.53 5.93 6.94 7.95 4.66 5.45 6.25 10:00 3.86 4.51 5.17 4.04 4.72 5.41 4.42 5.17 5.92 3.81 4.45 5.10 11:00 3.60 4.21 4.82 3.48 4.07 4.66 3.89 4.55 5.21 3.46 4.05 4.64 12:00 3.53 4.14 4.74 3.24 3.79 4.34 3.68 4.31 4.94 3.33 3.90 4.47 13:00 3.63 4.24 4.86 3.17 3.71 4.25 3.66 4.28 4.91 3.35 3.92 4.50 14:00 3.93 4.60 5.26 3.24 3.80 4.35 3.81 4.46 5.11 3.53 4.13 4.74 15:00 4.67 5.46 6.26 3.49 4.08 4.67 4.23 4.95 5.67 3.98 4.65 5.33 平均 3.96 4.63 5.30 3.75 4.39 5.03 4.23 4.95 5.67 3.73 4.36 5.00 3.2.2 光伏组件铺设高度对遮阴宽度的影响
为研究不同光伏组件铺设高度对遮阴宽度的影响,选择2.5、3.2、3.9 m的3种光伏组件铺设高度对遮阴宽度进行分析,默认光伏阵列的结构参数除光伏组件铺设高度外均相同。由表1可见,当光伏组件铺设高度越高时,南京、昆明、西安和长沙4地在任一时间段内遮阴宽度均越大,且下午遮阴宽度均值大于上午。无花果生育期内,当光伏组件铺设高度由2.5 m增高至3.9 m时,南京、昆明、西安和长沙4地遮阴宽度均值增幅分别为34.0%、34.0%、34.2%和34.1%。同时,以2.5 m的光伏组件铺设高度为例,无花果生育期内南京、昆明、西安和长沙4地的最大遮阴宽度占光伏阵列板间种植区域宽度的比例分别为34.6%、25.5%、36.9%和29.3%。
3.3 作物产量及光合特性分析
3.3.1 果实产量及其构成
光伏组件铺设高度对无花果产量及其构成的影响如表2所示。由表2可知,光伏农业系统内部各处理的产量均显著(P<0.05)低于露天对照,且光伏组件铺设高度对无花果产量及其构成指标的影响均达到显著水平(P<0.05)。与露天对照相比,H2.5、H3.2和H3.9的产量分别减少了19.9%、33.8%和48.9%。由此可见,无花果产量随着光伏组件铺设高度的增加而降低,H2.5较H3.2和H3.9分别增产13.9%和29.0%。在产量构成因素方面,各处理的单果质量和果数均显著(P<0.05)低于露天对照,且各处理间差异显著(P<0.05)。与露天对照相比,H2.5、H3.2和H3.9的单果质量分别降低了13.4%、22.6%和34.0%;果数分别减少了7.4%、14.3%和22.5%。由此可见,H2.5的无花果单果质量和果数等产量构成指标表现最优,而H3.9的各产量构成指标表现最差。
表 2 不同高度的光伏阵列对无花果果实产量及其构成的影响Table 2. PEffects of PV panels with different laying heights on yield and composition of fig fruit处理
Treatments单果质量
Single fruit weight/g平均单株果数
Average number of fruits
per plant果实产量
Fruit yield/(t·hm−2)CK 49.97±3.08a 135.20±4.32a 3.31±0.11a H2.5 43.27±4.14b 125.20±3.11b 2.65±0.07b H3.2 38.67±2.41c 115.80±4.96c 2.19±0.09c H3.9 32.99±3.72d 104.80±4.92d 1.69±0.08d 注:CK代表对照处理,同一列不同字母表示差异显著(P<0.05)。下同。 Note: CK represents the control treatment. Different letters in each column indicate significant differences at 0.05 level. The same as below. 3.3.2 光合特性
不同光伏组件铺设高度对光伏农业系统内部无花果叶片光合特性的测试结果如表3所示。
表 3 不同高度的光伏阵列对无花果叶片光合特性的影响Table 3. Effects of photovoltaic panels with different laying heights on photosynthetic characteristics of fig leaves处理
Treatments净光合速率
Net photosynthesis rate/
(μmol·m−2·s−1)气孔导度
Stomatol conductance/
(mol·m−2·s−1)蒸腾速率
Transperation rate/
(mmol·m−2·s−1)胞间二氧化碳
Intercellular carbon dioxide/
(μmol·m−2·s−1)CK 11.05±1.07a 14.62±2.11a 0.58±0.06a 343.47±15.64a H2.5 10.63±1.09a 13.68±2.28ab 0.53±0.07b 342.58±15.42a H3.2 9.94±0.58b 12.89±1.65b 0.51±0.03b 341.40±12.16a H3.9 8.49±0.93c 10.98±1.75c 0.37±0.05c 341.97±11.87a 由表3可知,在3种光伏组件铺设高度的光伏农业系统内部,无花果叶片的净光合速率、气孔导度和蒸腾速率均随着光伏组件铺设高度的增加而降低,且均显著(P<0.05)低于露天对照;而无花果叶片的胞间二氧化碳在各处理之间没有明显差异。与露天对照相比,H2.5、H3.2和H3.9的无花果叶片净光合速率分别降低了6.3%、11.7%和24.8%;气孔导度分别降低了9.2%、13.0%和36.6%;蒸腾速率分别降低了3.8%、10.1%和23.2%,且各处理间差异显著(P<0.05)。
4. 讨 论
4.1 光伏组件遮阴对光环境及遮阴宽度的影响
本研究发现不同光伏组件铺设高度下光伏阵列板间种植区域的采光率保持在55.4%~68.9%,是光伏阵列板下种植区域的2.1~3.3倍。魏来等[20]研究发现3排光伏组件的行间地面日平均太阳辐射强度比露天对照约弱18%,且日太阳辐射强度在12:00—14:00来回波动达到最大值。本研究观测的光伏农业系统内部的光环境变化规律与魏来等[20]的研究结果较为一致。然而魏来等[20]研究结果中的采光环境优于本研究结果,可能是由于其搭建的光伏阵列试验平台跨数较少,太阳辐射强度测点在一定程度上受到了光伏阵列外部侧光的影响。
在3种光伏组件铺设高度下,光伏阵列板间种植区域的平均太阳辐射强度和采光率随着光伏组件铺设高度的增加而降低,光伏阵列板下种植区域相反。结合光伏组件遮阴宽度的理论分析可以发现,在不同地理位置和光伏组件铺设高度下形成的遮阴宽度均会对光伏阵列板间种植区域的无花果生长造成影响。以南京地区为例,遮阴宽度最大值随着光伏组件铺设高度的增加而增大。在2.5 m的光伏组件铺设高度下,当遮阴宽度达到最大值时,阴影会覆盖无花果冠幅的18.9%;而在3.9 m的光伏组件铺设高度下,当遮阴宽度达到最大值时,阴影会覆盖无花果冠幅的29.9%。由此可见,光伏组件遮阴对光环境及遮阴宽度的影响具有一致性。
4.2 光伏组件遮阴对无花果产量及叶片光合特性的影响
在光伏阵列板间种植区域采光率为55.4%~68.9%的光伏农业系统内部,无花果树仍能完成正常的生命周期。适宜的光伏组件铺设高度能为无花果树提供一个较为理想的生长环境,从而实现光伏发电和农业生产的有效耦合。本研究中H2.5的无花果产量最高,达到了露天对照的80%,满足地方政府对该光伏农业项目产量的考核要求,这与H2.5拥有更好的光环境有关。由此可见,适宜的光伏组件铺设高度(2.5 m)能在保证正常完成生长结实的基础上,提升果树的结果数和单果质量,从而提升无花果的产量。结合试验光伏农业系统的无花果产量统计结果分析,H3.9的无花果产量比H2.5减产0.96 t/hm2。以一个20 MW的无花果光伏农业园区(约占地1.67 hm2/MW)为例,占地约33.3 hm2,可见H3.9无花果总产量将会比H2.5减少约32.0 t。以江苏省为例,仅2022年以来全省新增光伏农业市场化并网项目约1 388 MW(数据来源于江苏省发展与改革委员会官网),若不同光伏组件铺设高度下采用合理的农业生产模式,将极大改善农光耦合生产效果。
叶片是作物进行光合作用和呼吸作用等生理代谢活动的重要器官。遮阴胁迫情况下,作物净光合速率受到“气孔因素”[31-32]和“非气孔因素”[33]的限制。由试验数据可见,在光伏组件铺设高度逐渐增大的遮阴水平下,无花果叶片净光合速率和气孔导度显著降低(P <0.05),且光伏遮阴效应对胞间二氧化碳没有明显影响,说明净光合速率降低是由气孔因素和非气孔因素共同作用引起的,即可能是因为遮阴会减少无花果树叶片的气孔开张,降低水分的利用效率,从而影响光合作用的进行。本研究中,H2.5处理能够通过有效提高无花果叶片的净光合速率和气孔导度来缓解光合抑制。结合目前中国各地方发布的光伏组件铺设高度下限要求,说明2.5 m的光伏组件铺设高度更有利于保证无花果叶片的光合能力,更适于无花果树种植生产。
虽然中国的光伏农业模式在经济、社会和生态等方面取得了显著效益,成功解决了长期以来中国光伏发电产业存在的产能过剩等问题,带动了农业农村地区增收脱贫。但是现有的光伏农业系统在结构上和农业种植模式上还存在一系列问题,如在现有的结构下农作物产量低,难以实现可复制、可推广的农业生产技术体系等。未来中国光伏农业必然向提质增效方面发展,通过合理利用每一寸土地,最终提高光伏农业产出率。因为光伏阵列的板间种植区域是光伏农业系统中农田部分接受太阳能的最佳区域,所以在实际生产中需优先保障该区域的农业高效生产,针对不同的光伏组件铺设高度提出合适的农业生产方案。然而,影响光伏农业系统高效耦合生产的因素还有较多,诸如光伏阵列跨度、光伏组件铺设密度、作物种类和种植模式等,这些内容将在今后的研究中进一步丰富和深入。
5. 结 论
本文通过试验测试,量化了光伏组件遮阴对光伏农业系统内部光环境分布和农作物产量的影响。同时,通过太阳直射辐射理论计算分析了光伏组件遮阴形成的阴影宽度在一年中的变化规律,得到了如下结论:
1)研究发现,试验期间光伏阵列板间种植区域的采光率保持在55.4%~68.9%,是光伏阵列板下种植区域的2.1~3.3倍。在3种光伏组件铺设高度下,光伏阵列板间种植区域的平均太阳辐射强度和采光率随着光伏组件铺设高度的增加而降低,光伏阵列板下种植区域相反。
2)遮阴宽度受光伏组件宽度、光伏组件倾角、光伏组件铺设高度、太阳高度角和太阳方位角共5个参数的影响。在无花果生育期内(4—9月),光伏组件遮阴宽度随纬度、光伏组件铺设高度增大而增大。
3)光伏组件遮阴会显著降低无花果的产量,光伏组件铺设高度为2.5、3.2、3.9 m的产量较露天对照分别减少了19.9%、33.8%和48.9%,且叶片光合特性的测试结果验证了各处理间的产量差异。较低的光伏组件铺设高度能有效提高无花果叶片的净光合速率和气孔导度,从而减轻光伏组件遮阴导致的光合抑制。
研究发现,不同光伏组件铺设高度遮阴会严重影响光伏农业系统内部光环境的分布和作物的产量。基于此,建议如下:1)光伏农业生产上,在2.5 m等低高度的光伏阵列板间种植区域可种植无花果等同类果树;在较大高度的光伏阵列板间种植区域可种植中草药等喜荫作物,尽量避免因光伏组件铺设高度增大造成的较差光环境对农作物生长造成不利影响。2)光伏农业项目规划设计上,一方面应积极结合地方典型作物品种,做好光伏农业项目的规划设计;同时应积极开发、推广能一定程度上改善光伏阵列内部光环境的新型光伏农业阵列结构,以保证农作物的正常生长。
-
![]()
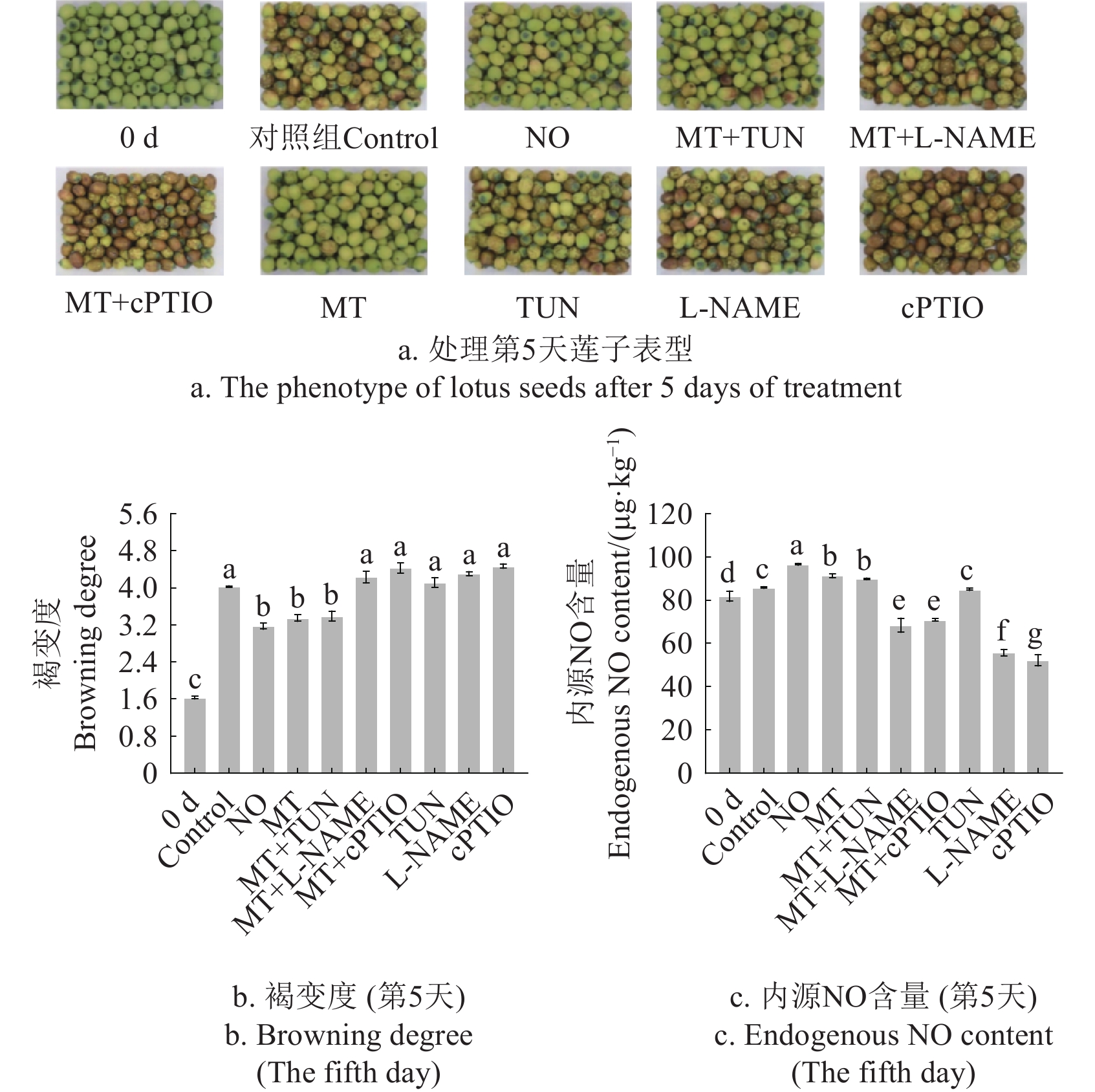
图 1 不同处理对采后莲子褐变和NO含量的影响
注:Control为蒸馏水处理组,NO为一氧化氮处理组,MT为褪黑素处理组,TUN为硝酸还原酶抑制剂处理组,L-NAME为一氧化氮合酶抑制剂处理组,cPTIO为一氧化氮清除剂处理组,MT+TUN为MT结合TUN处理组,MT+L-NAME为MT结合L-NAME处理组,MT+cPTIO为MT结合cPTIO处理组。不同字母表示处理间具有显著性差异( P < 0.05),下同。
Figure 1. Effects of different treatments on the browning and NO content of postharvest lotus seeds
Note: The same as below;Control indicates distilled water treatment, NO indicates nitric oxide treatment, MT indicates melatonin treatment, TUN indicates tungstate treatment, L-NAME indicates nomega-nitro-L-arginine methyl ester treatment, cPTIO indicates carboxy-PTIO treatment, MT+TUN indicates MT combined with TUN treatment, MT+L-NAME indicates MT combined with L-NAME treatment, MT+cPTIO indicates MT combined with cPTIO treatment. Different letters at the same time point indicate significant differences (P < 0.05). Same as below.
![]()
图 2 不同处理对采后莲子表型和莲子皮褐变度的影响
Figure 2. Effects of different treatments on phenotype of postharvest lotus seeds and browning degree of postharvest lotus seed coats
![]()
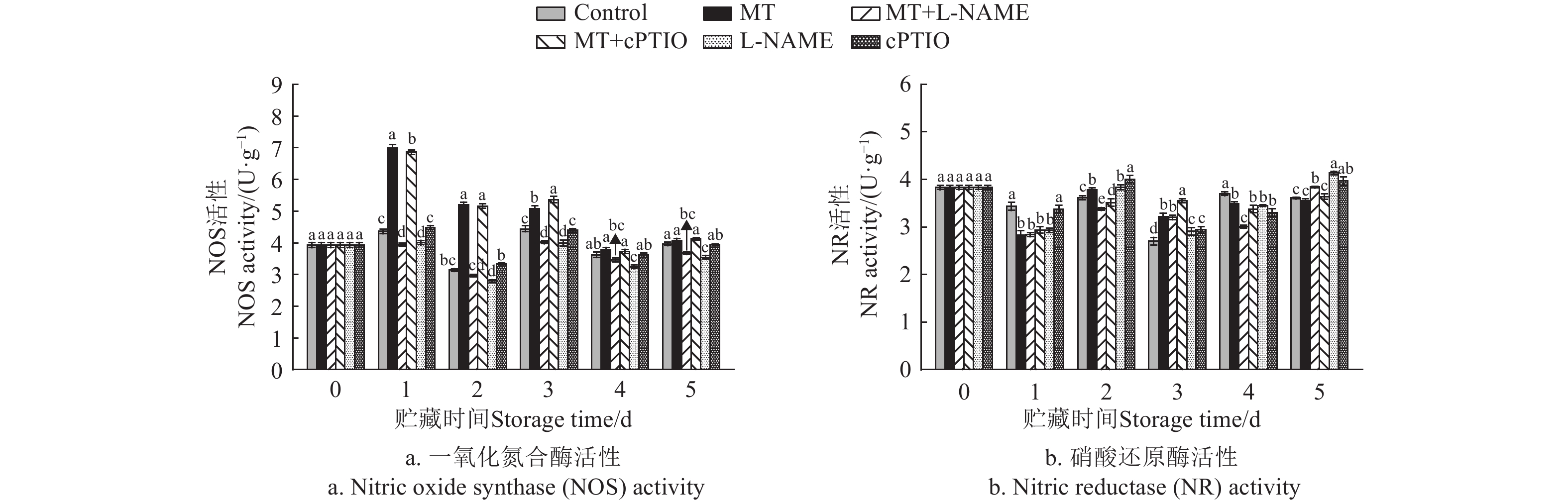
图 3 不同处理对采后莲子NOS和NR活性的影响
Figure 3. Effects of different treatments on NOS and NR activities of postharvest lotus seeds
![]()
图 4 不同处理对采后莲子L-精氨酸和瓜氨酸含量的影响
Figure 4. Effects of different treatments on L-arginine and citrulline contents of postharvest lotus seeds
![]()
图 5 不同处理对采后莲子总酚和儿茶素含量的影响
Figure 5. Effects of different treatments on total phenol and catechin contents of postharvest lotus seeds
![]()
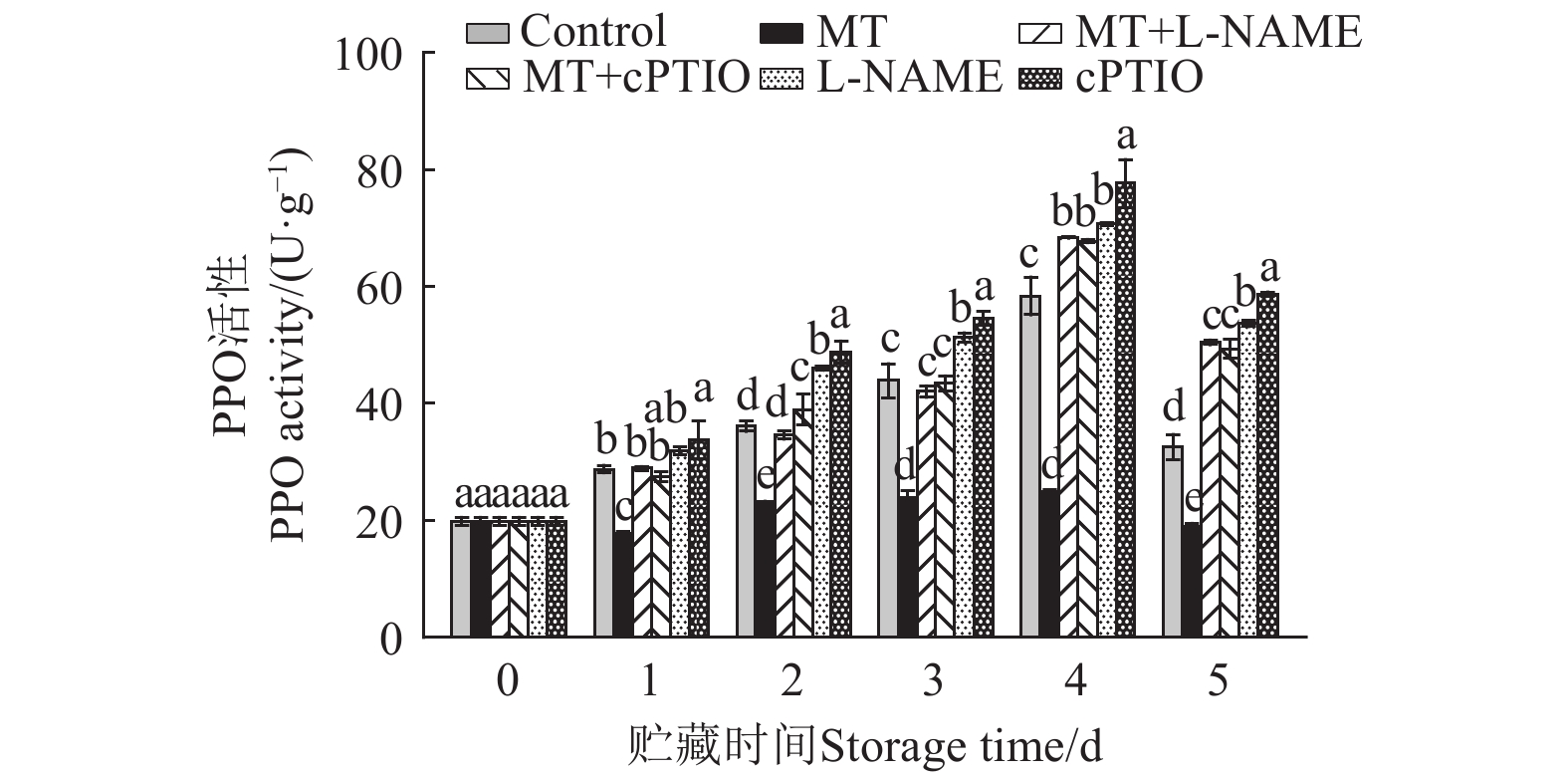
图 6 不同处理对采后莲子PPO活性的影响
Figure 6. Effects of different treatments on PPO activity of postharvest lotus seeds
![]()
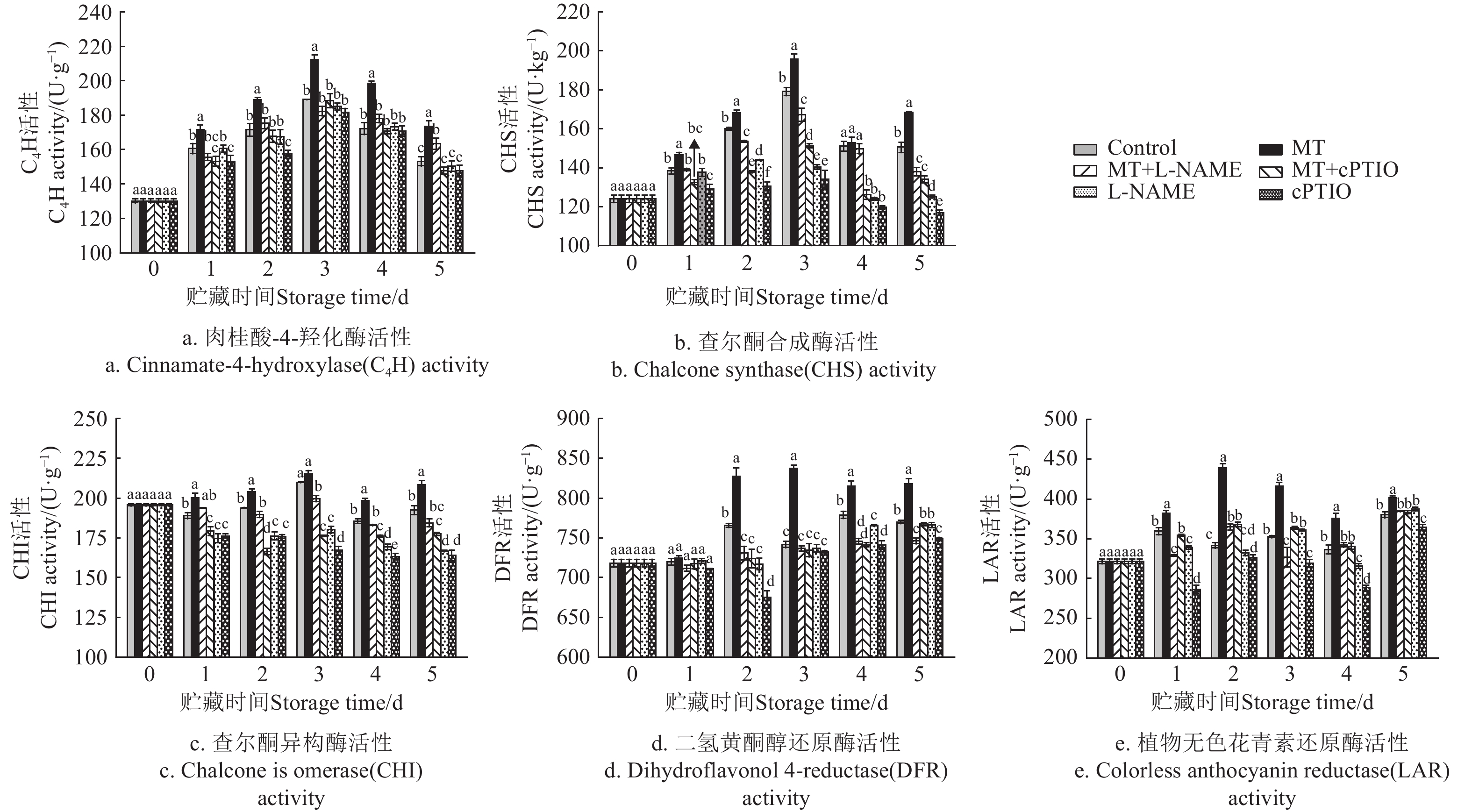
图 7 不同处理对采后莲子C4H、CHS、CHI、DFR和LAR活性的影响
Figure 7. Effects of different treatments on C4H, CHS, CHI, DFR and LAR activities of postharvest lotus seeds
![]()
图 8 采后鲜莲子对外源MT处理的响应模式图
注:NOS表示为一氧化氮合酶。
Figure 8. Possible model of postharvest lotus seeds response to MT treatment
Note: NOS indicates nitric oxide synthase.
表 1 鲜莲子中的酚类物质及其含量
Table 1 The phenolic compounds and their contents in fresh lotus seeds
酚类物质
Phenolic compounds化学式
Chemical formula分子量
Neutral mass保留时间
Retention time/min母离子
Parent ion [M-H]−/
(m·z−1)碎片离子
Fragments MS2/(m·z−1)含量
Content/
(mg·kg−1)儿茶素Catechin C15H14O6 290.05 9.06 289.05 245.06,203.07,137.01,123.03,109.03 51.46 表儿茶素Epicatechin C15H14O6 290.05 10.99 289.05 245.06,203.07,137.01,123.03,109.03 5.94 芦丁Rutin C27H30O16 610.10 17.18 609.09 301.01,300.03 0.77 金丝桃苷Hyperin C21H20O12 464.06 17.43 463.05 300.00,179.00 3.70 异槲皮苷Isoquercitrin C21H20O12 464.06 19.91 463.08 300.00,179.00 0.92 槲皮素Quercetin C15H10O7 302.02 23.80 301.01 178.98,150.99 2.01 没食子酸Gallic acid C7H6O5 170.00 3.40 169.00 125.01 2.71 肉桂酸Cinnamic acid C9H8O2 148.06 7.21 147.06 85.02 5.01 水杨酸Salicylic acid C7H6O3 138.02 9.22 137.01 93.03 1.39 p-香豆酸P-coumaric acid C9H8O3 164.04 13.41 163.04 119.04 0.24 阿魏酸Ferulic acid C6H10O7 194.04 14.44 193.03 178.01,149.05,134.03 0.91 芥子酸Sinapic acid C11H12O5 224.05 14.56 223.04 208.02,193.00,149.01,121.02 3.73 鞣花酸Ellagitannin C14H6O8 302.01 23.84 301.01 178.98,150.99,121.02,107.00 6.34 总含量Total content 85.13  下载: 导出CSV
下载: 导出CSV
-
[1] 辛佳,孙恒,刘娟,等. 莲黄酮类化合物鉴定、功能与应用的研究进展[J]. 植物科学学报,2023,41(3):400-410. doi: 10.11913/PSJ.2095-0837.22194 XIN Jia, SUN Heng, LIU Juan, et al. Advances in the identification, function, and application of flavonoids in Nelumbo[J]. Plant Science Journal, 2023, 41(3): 400-410. (in Chinese with English abstract) doi: 10.11913/PSJ.2095-0837.22194
[2] YANG Y A, LIU R L, HAN Y H, et al. Critical taste substances and regulatory pathways of fresh lotus seed pulps at different ripeness stages[J]. Postharvest Biology and Technology, 2023, 205: 112522. doi: 10.1016/j.postharvbio.2023.112522
[3] 高建晓,王毓宁,李鹏霞,等. 漆蜡涂膜对鲜莲蓬采后褐变的影响[J]. 食品科学,2016,37(18):275-282. doi: 10.7506/spkx1002-6630-201618044 GAO Jianxiao, WANG Yuning, LI Pengxia, et al. Effects of lacquer wax coating on postharvest browning of fresh lotus pods[J]. Food Science, 2016, 37(18): 275-282. (in Chinese with English abstract) doi: 10.7506/spkx1002-6630-201618044
[4] LI P X, HU H L, LUO S F, et al. Shelf life extension of fresh lotus pods and seeds (Nelumbo nucifera Gaertn. ) in response to treatments with 1-MCP and lacquer wax[J]. Postharvest Biology and Technology, 2017, 125: 140-149. doi: 10.1016/j.postharvbio.2016.10.004
[5] 李梦蝶,谢湘汝,李娟娟. 鲜莲子采后保鲜的研究进展[J]. 湖北工程学院学报,2022,42(6):61-67. doi: 10.3969/j.issn.2095-4824.2022.06.012 LI Mengdie, XIE Xiangru, LI Juanjuan. Advance in the research on the preservation of fresh lotus seeds[J]. Journal of Hubei Engineering University, 2022, 42(6): 61-67. (in Chinese with English abstract) doi: 10.3969/j.issn.2095-4824.2022.06.012
[6] JIANG Y M, DUAN X W, QU H, et al. Browning: enzymatic browning[J]. Encyclopedia of Food and Health, 2016: 508–514.
[7] 李京芳. 莲藕酶促褐变及功能成分变化对降脂、降胆固醇的作用和机理探讨[D]. 南昌:南昌大学,2023:67-68. LI Jingfang. Effects of the Enzymatic Browning of Lotus Root and the Transformation of Functional Components on Regulating Lipid and Cholesterol Metabolism [D]. Nanchang: Nanchang University, 2023: 67-68. (in Chinese with English abstract)
[8] 薛建新,王凯,李泽珍,等. 采后褪黑素处理对鲜切花椰菜保鲜品质及货架期的影响[J]. 农业工程学报,2021,37(13):273-283. XUE Jianxin, WANG Kai, LI Zezhen, et al. Influences of post-harvest melatonin treatment on preservation quality and shelf life of fresh-cut cauliflower[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2021, 37(13): 273-283. (in English with Chinese abstract)
[9] LI P P, ZHANG R H, ZHOU H D. Melatonin delays softening of postharvest pepper fruits (Capsicum annuum L.) by regulating cell wall degradation, membrane stability and antioxidant systems[J]. Postharvest Biology and Technology, 2024, 212: 112852. doi: 10.1016/j.postharvbio.2024.112852
[10] CAI S Y, ZHANG Z Q, WANG J L. Effect of exogenous melatonin on postharvest storage quality of passion fruit through antioxidant metabolism[J]. LWT - Food Science and Technology, 2024, 194: 11583.
[11] WANG Z Q, PI H L, SHAN S S, et al. Melatonin enhanced chilling tolerance and alleviated peel browning of banana fruit under low temperature storage[J]. Postharvest Biology and Technology, 2021, 179: 111571. doi: 10.1016/j.postharvbio.2021.111571
[12] ONIK J C, SU C W, LI A, et al. Melatonin treatment reduces ethylene production and maintains fruit quality in apple during postharvest storage[J]. Food Chemistry, 2020, 337: 127753.
[13] 张亚琳,王慧慧,芦玉佳. 外源褪黑素对杏果实采后黑斑病的调控作用[J]. 中国食品学报,2024,24(3):201-209. ZHANG Yalin, WANG Huihui, LU Yujia, et al. Regulation of postharvest black spot disease in apricot fruit by exogenous melatonin[J]. Journal of Chinese Institute of Food Science and Technology, 2024, 24(3): 201-209. (in Chinese with English abstract)
[14] 董小盼,汤静,丁娇,等. 褪黑素处理对桃果实采后软腐病的影响及其机理[J]. 食品科学,2024,45(11):243-249. doi: 10.7506/spkx1002-6630-20230918-160 DONG Xiaopan, TANG Jing, DING Jiao, et al. Effects of melatonin treatment on rhizopus rot in postharvest peach fruit and possible mechanism[J]. Food Science, 2024, 45(11): 243-249. (in Chinese with English abstract) doi: 10.7506/spkx1002-6630-20230918-160
[15] SONG L J, ZHANG W W, QIANG L. Melatonin alleviates chilling injury and maintains postharvest quality by enhancing antioxidant capacity and inhibiting cell wall degradation in cold-stored eggplant fruit[J]. Postharvest Biology and Technology, 2022, 194: 112092. doi: 10.1016/j.postharvbio.2022.112092
[16] 董萤飞. 独脚金内脂对采后草莓一氧化氮、硫化氢以及苯丙烷代谢的影响[D]. 泰安:山东农业大学,2020:3-4. DONG Yingfei. Effects of Strigolactone on NO, H2S and Phenylpropane Metabolism in Postharvest Strawberry Fruits. [D]. Taian: Shandong Agricultural University, 2020: 3-4. (in Chinese with English abstract)
[17] POLS S, VAN DE POEL B, HERTOG M L A T M, et al. The regulatory role of nitric oxide and its significance for future postharvest applications[J]. Postharvest Biology and Technology, 2022, 188: 111869. doi: 10.1016/j.postharvbio.2022.111869
[18] 陈燕,张健,魏佳,等. 一氧化氮熏蒸抑制干制灰枣黑曲霉病及贮藏品质保持[J]. 农业工程学报,2019,35(12):297-303. doi: 10.11975/j.issn.1002-6819.2019.12.036 CHEN Yan, ZHANG Jian, WEI Jia, et al. Nitric oxide fumigation inhibiting Aspergillus niger disease and maintaining storage quality of dried ash jujube[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2019, 35(12): 297-303. (in Chinese with English abstract) doi: 10.11975/j.issn.1002-6819.2019.12.036
[19] LIU J L, YANG J, ZHANG H Q, et al. Melatonin inhibits ethylene synthesis via nitric oxide regulation to delay postharvest senescence in pears[J]. Journal of Agricultural and Food Chemistry, 2019, 67: 2279-2288. doi: 10.1021/acs.jafc.8b06580
[20] AGHDAM M S, LUO Z, JANNATIZADEH A, et al. Employing exogenous melatonin applying confers chilling tolerance in tomato fruits by upregulating ZAT2/6/12 giving rise to promoting endogenous polyamines, proline, and nitric oxide accumulation by triggering arginine pathway activity[J]. Food Chemistry, 2019, 275: 549-556. doi: 10.1016/j.foodchem.2018.09.157
[21] SUN L, LUO S F, HU H L, et al. Melatonin promotes the normal cellular mitochondrial function of lotus seeds through stimulating nitric oxide production[J]. Postharvest Biology and Technology, 2022, 185: 111814.
[22] SUN H, LI J, SONG W, et al. Nitric oxide generated by nitrate reductase increases nitrogen uptake capacity by inducing lateral root formation and inorganic nitrogen uptake under partial nitrate nutrition in rice[J]. Journal of Experimental Botany, 2015, 66(9): 2449-2459. doi: 10.1093/jxb/erv030
[23] ZAFARI S, SHAEIFI M, CHASHMI N. Nitric oxide production shifts metabolic pathways toward lignification to alleviate Pb stress in Prosopis farcta[J]. Environmental and Experimental Botany, 2017, 141: 41-49. doi: 10.1016/j.envexpbot.2017.06.011
[24] ZHANG Q, ZHANG L, GENG B, et al. Interactive effects of abscisic acid and nitric oxide on chilling resistance and active oxygen metabolism in peach fruit during cold storage[J]. Journal of the Science of Food and Agriculture, 2019, 99(7): 3367-3380. doi: 10.1002/jsfa.9554
[25] DONG M, QIN L, XUE J, et al. Simultaneous quantification of free amino acids and 5'-nucleotides in shiitake mushrooms by stable isotope labeling-LC-MS/MS analysis[J]. Food Chemistry, 2018, 268: 57-65. doi: 10.1016/j.foodchem.2018.06.054
[26] CHEN S, XIE R, LI J, et al. Alteration on phenolic acids and the appearance of lotus (Nelumbo nucifera Gaertn) seeds dealt with antistaling agents during storage[J]. International Journal of Food Properties, 2018, 21(1): 1481-1494.
[27] 贾文婷,李文绮,吴洪斌. 不同前处理联合压差闪蒸干燥对红枣脆片品质的影响[J]. 农业工程学报,2024,40(2):116-123. JIA Wenting, LI Wenqi, WU Hongbin. Effects of different pretreatments combined with instant controlled pressure drop drying on the quality of jujube slices[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2024, 40(2): 116-123. (in Chinese with English abstract)
[28] SHU P, MIN D, AI W, et al. L-Arginine treatment attenuates postharvest decay and maintains quality of strawberry fruit by promoting nitric oxide synthase pathway[J]. Postharvest Biology and Technology, 2020, 168: 111253. doi: 10.1016/j.postharvbio.2020.111253
[29] LI J, DENG Z, HE Y, et al. Differential specificities of polyphenol oxidase from lotus seeds (Nelumbo nucifera Gaertn.) toward stereoisomers, (−)-epicatechin and (+)-catechin: Insights from comparative molecular docking studies[J]. LWT - Food Science and Technology, 2021, 148: 111728. doi: 10.1016/j.lwt.2021.111728
[30] BAO Z Y, ZHOU Q H, YU Y, et al. Melatonin treatment induces DNA methylation to alleviate chilling induced-browning in cold stored peach fruit[J]. Postharvest Biology and Technology, 2024, 208: 112686. doi: 10.1016/j.postharvbio.2023.112686
[31] WANG Y, GAO L, WANG Z, et al. Light-induced expression of genes involved in phenylpropanoid biosynthetic pathways in callus of tea (Camellia sinensis L. O. Kuntze)[J]. Scientia Horticulturae, 2012, 133: 72-83. doi: 10.1016/j.scienta.2011.10.017
[32] 王甜. 外源褪黑素对荔枝采后褐变和病害的控制作用[D]. 海口:海南大学,2020:35-36. WANG Tian. Effect of Exogenous Melatonin on Postharvest Browning and Disease in Harvested Litchi Fruit[D]. Haikou: Hainan University, 2020: 35-36. (in Chinese with English abstract)
[33] 陈力维,令阳,邓丽莉,等. L-半胱氨酸处理对采后青脆李果实苯丙烷代谢的影响[J]. 农业工程学报,2020,36(13):257-263. doi: 10.11975/j.issn.1002-6819.2020.13.030 CHEN Liwei, LIING Yang, DENG Lili, et al. Effects of L-cysteine treatment on phenylpropanoid metabolism of postharvest “Qingcui” plum fruit[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2020, 36(13): 257-263. (in Chinese with English abstract) doi: 10.11975/j.issn.1002-6819.2020.13.030
[34] WANG M, LI Y H, LI C Y, et al. Melatonin induces resistance against Penicillium expansum in apple fruit through enhancing phenylpropanoid metabolism[J]. Physiological and Molecular Plant Pathology, 2023, 127: 102082. doi: 10.1016/j.pmpp.2023.102082
[35] WANG Q H, HAN Y, YANG R R. Melatonin facilitates the deposition of suberin polyphenolic and lignin at wounds of potato tubers by mediating nitric oxide and reactive oxygen species[J]. Postharvest Biology and Technology, 2023, 198: 112270. doi: 10.1016/j.postharvbio.2023.112270









计量
- 文章访问数: 0
- HTML全文浏览量: 0
- PDF下载量: 0





